独脚金内酯(strigolactone,SL)最初是从分析玉米根系中植物分泌的一种刺激恶性寄生杂草独角金(Striga spp.)种子萌发的信号物质时发现的通过类胡萝卜素途径合成的倍半萜类化合物,因此被称为独脚金内酯。后来进一步发现其具有多种生理功能,近年被证明为一种植物中普遍存在的新型植物激素。
独脚金内酯并不是指某种具体的植物激素,而是几种植物激素的统称。其包括:独脚金醇、高粱内酯、列当醇和人工合成的类似物GR24、GR6、GR7等。根据其化学结构,SLs可分为典型(Canonical SLs) 和非典型独脚金内酯(Non-canonical SLs)两类。
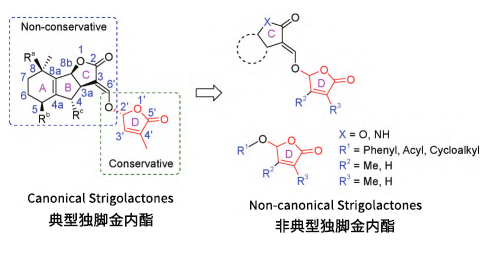
典型的SLs:具有相似的倍半萜碳骨架结构,可分为独脚金醇(Strigol)和列当醇(Orobanchol),它们均由一个三环(ABC环)内酯通过一个烯醇醚桥与5-羟基呋喃酮(D环)以醚键连接而成。
研究发现,烯醇醚桥和D环是其保持SL样生物活性的保守结构,D环C-2'的R立体构型是其保持强效发芽刺激活性的重要结构特征。在天然分离物的A、B环上发现了烷基、羟基或乙酰氧基等不同官能团修饰,构成了天然SLs的结构多样性。目前,已有30多种天然的独脚金内酯从棉花、水稻、红三叶草、豇豆和烟草等植物的根系分泌物中分离得到,并作为寄生杂草种子独脚金、小列当和分支列当的萌发刺激物活丛植菌根真菌的菌丝分支诱导物。
非典型的SLs:在研究独脚金内酯系列代谢产物的过程中发现的一类衍生物,其特点是含有保守的烯醇醚键和D环结构,但经典的倍半萜ABC环结构被多样化骨架的脂肪链所取代。
独脚金内酯的合成途径是以类胡萝卜素为前体,经脱氧、氧化等过程后生成独脚金醇,再由独脚金醇转化成具有生物活性的独脚金内酯物质。目前,已从拟南芥、水稻和豌豆等植物突变体中鉴定分离出和独脚金内酯代谢相关的4个酶类,分别为类胡萝卜素异构酶D27(β-carotenoid isomerase)、类胡萝卜素裂解双加氧酶7(carotenoid cleavagedioxygenase7,CCD7)、类胡萝卜素裂解双加氧酶8(carotenoid cleavagedioxygenase8,CCD8)以及细胞色素P450单加氧酶(cytochrome P450 monooxygen-ase,P450)。
合成过程:
目前SLs生物合成途径已基本明晰,其合成过程发生在质体和细胞质中。质体中的合成过程涉及异构酶D27、CCD7和CCD8:D27首先将all-trans-β-carotene异构化为9-cis-β-carotene;CCD7裂解9-cis-β-carotene的C9′-C10′的双键,使其生成9-cis-β-apo-10′-carotenal;CCD8进一步氧化裂解生成carlactone(CL)。随后CL转移至细胞质中,被CYP711A亚家族的MAX1(MOREAXILLARYGROWTH1)氧化生成carlactonoicacid(CLA);CLA被催化生成不同结构的SLs。
在水稻中,CLA被OsCYP711A2催化生成4-deoxyorobanchol;在玉米、番茄中,CLA分别被ZmMAX1b、CYP722C催化生成orobanchol;在棉花中,CLA被CYP722C催化生成5-deoxystrigol。
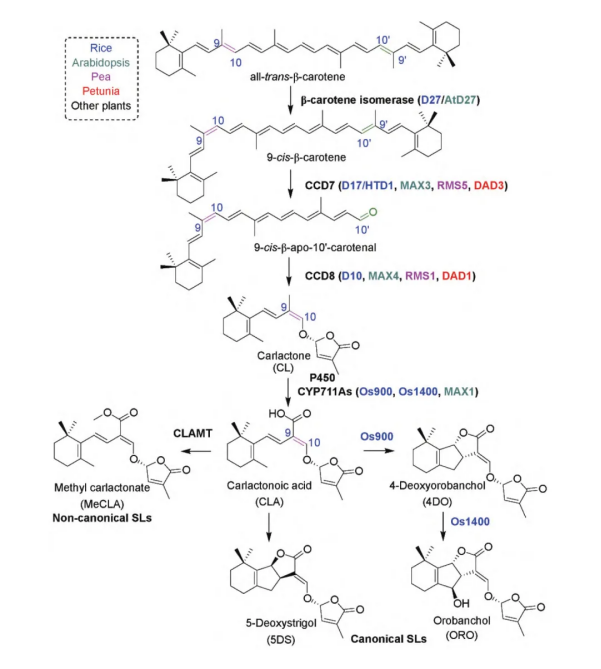
SLs在不同植物中的生物合成途径
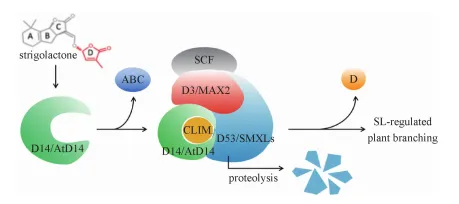
目前已鉴定的植物 SLs 信号转导相关基因包括
①D3(MAX2/RMS4)编码F-box蛋白,参与蛋白泛素化降解过程,突变体材料包括水稻d3、拟南芥max2、豌豆rms4 等;②D14(AtD14)编码α/β折叠水解酶超家族蛋白,突变体材料包括水稻d14、拟南芥Atd14 等;
③D53(SMXL6/7/8)编码SCF-E3泛素连接酶复合物的底物蛋白(行使 SLs 信号抑制因子),突变体材料包括水稻d53、拟南芥smxl6/7/8 等。
信号转导过程:根据对拟南芥、水稻SLs的信号转导机制研究发现 D14 是 SLs 的受体,其具有酶和受体的双重功能。SLs与D14蛋白结合,被后者水解成 ABC环与 D环,ABC环被释放,而D环留在D14结合口袋并形成中间体分子 CLIM ( covalently linked intermediate molecule),CLIM 与D14 催化中心共价结合,导致 D14 发生显著的构象变化并形成新的相互作用表面。随后,D14CLIM招募D3及SCF复合体,进一步形成SCFD14复合体,SCFD14通过其D3蛋白特异性识别结合D53,然后经由泛素结合酶E2介导将D53多泛素化修饰,修饰后的D53被26S蛋白酶体降解,从而解除了 D53对SLs信号通路的抑制,促使下游基因转录,实现 SLs信号转导。
独脚金内酯的生物功能
独脚金内酯不仅在植物根际起着信号调控的作用,还在植物的生长发育中也发挥着重要的作用。具体如下:
刺激寄生植物种子萌发:
植物根部分泌的 SLs能够促进寄生植物如列当、独脚金等的萌发。
促进丛枝菌根真菌菌丝分枝:
SLs通过促进菌丝分枝,有利于丛枝菌根真菌与植物有益共生关系的建立。
响应逆境胁迫:
SLs 响应干旱和盐胁迫,在磷酸盐和硝酸盐不足时,植物产生和释放更多的SLs。
抑制植物侧枝生长:
SLs能抑制侧芽生长以调节植物分枝,它在植物根和茎中合成、通过木质部向上运输或在茎局部直接抑制分枝。作为生长素下游因子,SLs通过抑制顶端优势的解除,从而调控侧枝的生长发育。
独脚金内酯与其他激素的交互作用:
SLs与脱落酸(ABA)、生长素(IAA)和细胞分裂素(CK)等激素存在交互作用,可协同调控植物的分支、根系发育等。